
We use cookies to ensure our website works properly and to personalise your experience. Cookies policy
Department of Pharmacology, Siddhant College of Pharmacy, Maval, Pune- 412109
Depression is a global health challenge with significant social and economic impact, and current pharmacological treatments are often limited by delayed onset, partial response, and adverse effects. This has prompted interest in herbal alternatives with multi target actions and better tolerability. Carissa spinarum Linn., a thorny shrub of the Apocynaceae family, has long been used in African, Asian, and Indian traditional medicine for its cardiotonic, anti inflammatory, and neuroactive properties. Phytochemical analyses of its root bark reveal alkaloids, flavonoids, triterpenoids, and sterols, compounds known to modulate serotonergic and noradrenergic pathways. Preclinical studies in Wistar rats demonstrate dose dependent antidepressant like activity, with behavioral improvements in Tail Suspension Tests, and neurochemical assays confirming enhanced serotonin and norepinephrine levels. Safety evaluations indicate tolerability at therapeutic doses, though caution is advised in reproductive contexts. These findings support C. spinarum as a promising candidate for integration into complementary medicine and future clinical validation.
Depression is one of the most prevalent neuropsychiatric disorders, affecting over 300 million individuals globally and contributing significantly to disability-adjusted life years (DALYs) (1). The World Health Organization (WHO) recognizes depression as a leading cause of global disease burden, with projections suggesting it may become the foremost contributor to morbidity by 2030 (2). Clinical manifestations include persistent sadness, loss of interest, cognitive dysfunction, and somatic symptoms, often leading to impaired social and occupational functioning (3). Despite advances in neuroscience, the etiology of depression remains multifactorial, involving genetic predisposition, environmental stressors, and neurochemical imbalances (4). Current pharmacological interventions primarily target monoaminergic neurotransmission, including selective serotonin reuptake inhibitors (SSRIs), serotonin-norepinephrine reuptake inhibitors (SNRIs), tricyclic antidepressants (TCAs), and monoamine oxidase inhibitors (MAOIs) (5). While effective in many patients, these drugs are limited by delayed onset of action (typically 2–6 weeks), partial or non-response in up to 30% of cases, and adverse effects such as sexual dysfunction, weight gain, and insomnia (6). Moreover, treatment-resistant depression remains a major clinical challenge, necessitating exploration of novel therapeutic strategies (7).
The serotonergic (5-HT) and noradrenergic (NA) systems are central to the pathophysiology of depression. Dysregulation of serotonin transporters (SERT) and norepinephrine transporters (NET) leads to impaired neurotransmission, contributing to mood disturbances (8). SSRIs act by inhibiting SERT, thereby increasing synaptic serotonin, while SNRIs block both SERT and NET, enhancing serotonergic and noradrenergic signaling (9). Preclinical rodent models have consistently demonstrated that modulation of these pathways produces robust antidepressant-like effects, validating their translational relevance (10).
Herbal remedies have been integral to traditional medicine systems across cultures, including Ayurveda, Traditional Chinese Medicine (TCM), and African ethnomedicine. Plants such as Hypericum perforatum (St. John’s Wort) have demonstrated clinical efficacy in mild-to-moderate depression, highlighting the therapeutic potential of phytochemicals (11). Ethnopharmacological surveys reveal that communities have long relied on plant extracts for mood regulation, often with fewer side effects compared to synthetic drugs (12). Phytochemicals such as flavonoids, alkaloids, and terpenoids exert neuroactive effects by modulating monoamine transporters, receptors, and enzymes. For instance, quercetin inhibits monoamine oxidase (MAO), enhancing serotonin and norepinephrine levels (13). Alkaloids like harmine interact with serotonergic receptors, while triterpenoids such as lupeol exhibit neuroprotective and anti-inflammatory properties (14). These multi-target actions suggest that herbal compounds may provide broader therapeutic coverage than single-target synthetic drugs (15).
Herbal medicines offer several advantages:
However, challenges include variability in phytochemical composition, lack of standardization, and limited clinical trials (17).
Carissa spinarum Linn. (Apocynaceae), commonly known as conkerberry, is a thorny shrub distributed across Africa, Asia, and India. It thrives in semi-arid regions and is recognized for its medicinal root bark (18). Ethnomedicinal records indicate its use as a cardiotonic, anti-inflammatory, and neuroactive agent. In African traditional medicine, root bark decoctions are employed for fever, pain, and nervous disorders, while in Ayurveda, it is used for gastrointestinal and cardiovascular ailments (19). Phytochemical analyses reveal the presence of alkaloids (carissin), flavonoids (quercetin), triterpenoids (lupeol), and sterols (β-sitosterol). These compounds are implicated in CNS modulation, antioxidant activity, and neuroprotection (20,21).
Wistar rats are widely used in neuropsychiatric research due to their genetic stability, reproducibility, and behavioral responsiveness. They provide reliable models for evaluating antidepressant-like activity (22).
Behavioral Paradigms
Neurochemical Assays
Quantification of serotonin and norepinephrine levels in brain tissue, along with receptor binding assays, provides mechanistic insights into antidepressant activity (24).
Mechanistic Pathways
Serotonergic Modulation
Extracts may act via 5-HT1A receptor agonism, 5-HT2 receptor modulation, and inhibition of SERT, thereby enhancing serotonergic transmission (25).
Noradrenergic Modulation
Noradrenergic effects include inhibition of NET and modulation of α2-adrenoceptors, leading to increased synaptic norepinephrine (26).
Cross-talk Between Monoaminergic Systems
Evidence suggests synergistic interactions between serotonergic and noradrenergic pathways, with dual modulation producing superior antidepressant effects (27).
Given the global burden of depression and limitations of current pharmacotherapy, evaluating the antidepressant potential of C. spinarum root bark extract in validated animal models provides a scientific basis for its integration into complementary medicine. This study aims to elucidate serotonergic and noradrenergic involvement, thereby contributing to the rational development of herbal-based psychotherapeutics (28–30).
Table 1. Comparative Features of Conventional vs Herbal Antidepressants
|
Parameter |
Conventional Antidepressants |
Herbal Extracts (e.g., C. spinarum) |
|
Primary mechanism |
Monoamine reuptake inhibition (SSRIs, SNRIs) |
Multi-target modulation (5-HT, NA, GABA) |
|
Onset of action |
2–6 weeks |
Potentially faster (preclinical evidence) |
|
Side effects |
Sexual dysfunction, weight gain, insomnia |
Generally fewer, but require toxicity validation |
|
Cultural acceptance |
Moderate |
High in traditional medicine systems |
|
Research status |
Extensive clinical trials |
Limited but growing preclinical evidence |
Table 2. Phytochemicals in Carissa spinarum and Neuroactive Potential
|
Compound |
Class |
Reported Activity |
Reference |
|
Carissin |
Alkaloid |
CNS stimulation, antidepressant-like |
(20) |
|
Lupeol |
Triterpenoid |
Anti-inflammatory, neuroprotective |
(21) |
|
Quercetin |
Flavonoid |
Antioxidant, MAO inhibition |
(14) |
|
β-sitosterol |
Sterol |
Neuroprotective, adaptogenic |
(15) |
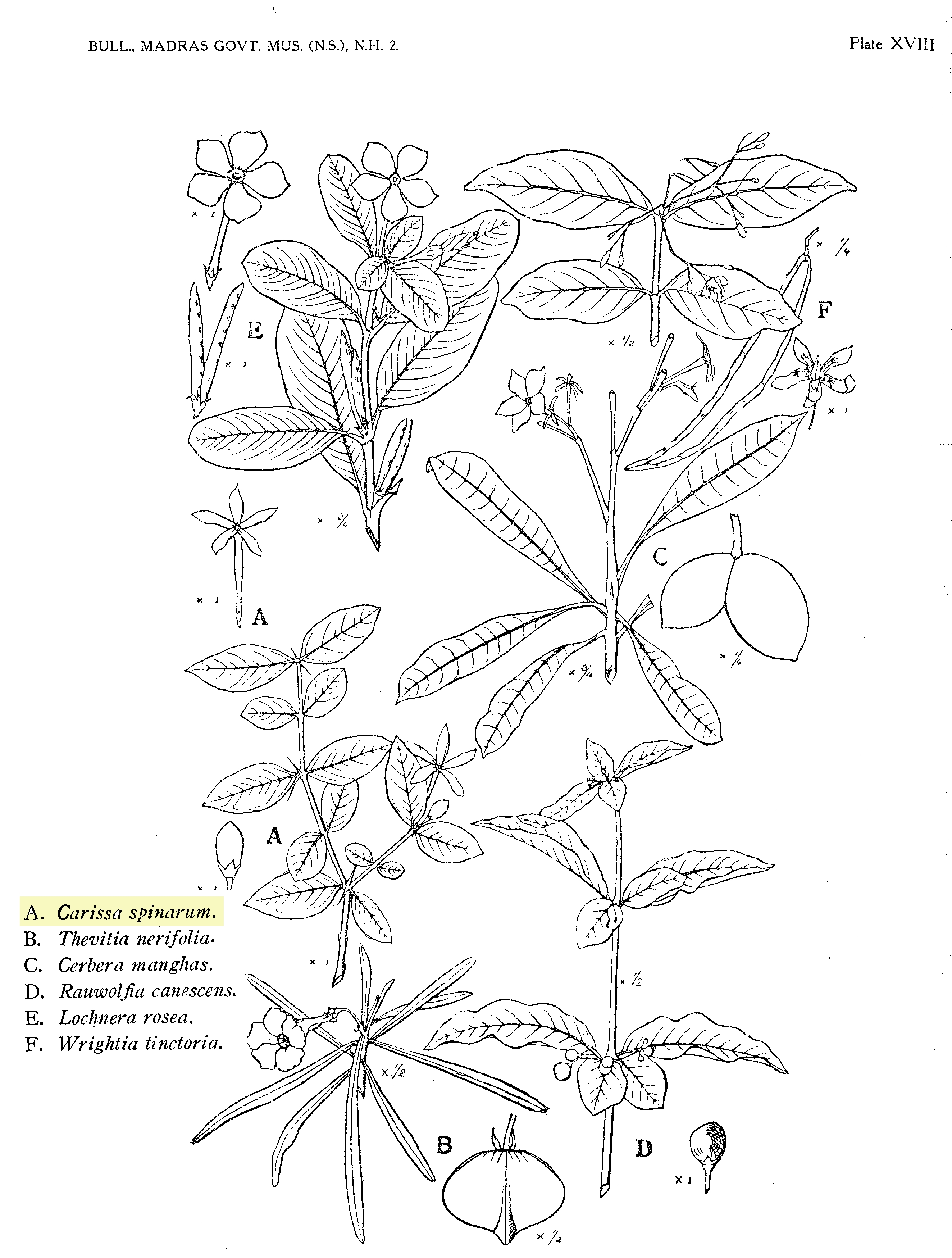
Figure 1: Botanical illustration of Carissa spinarum root bark .
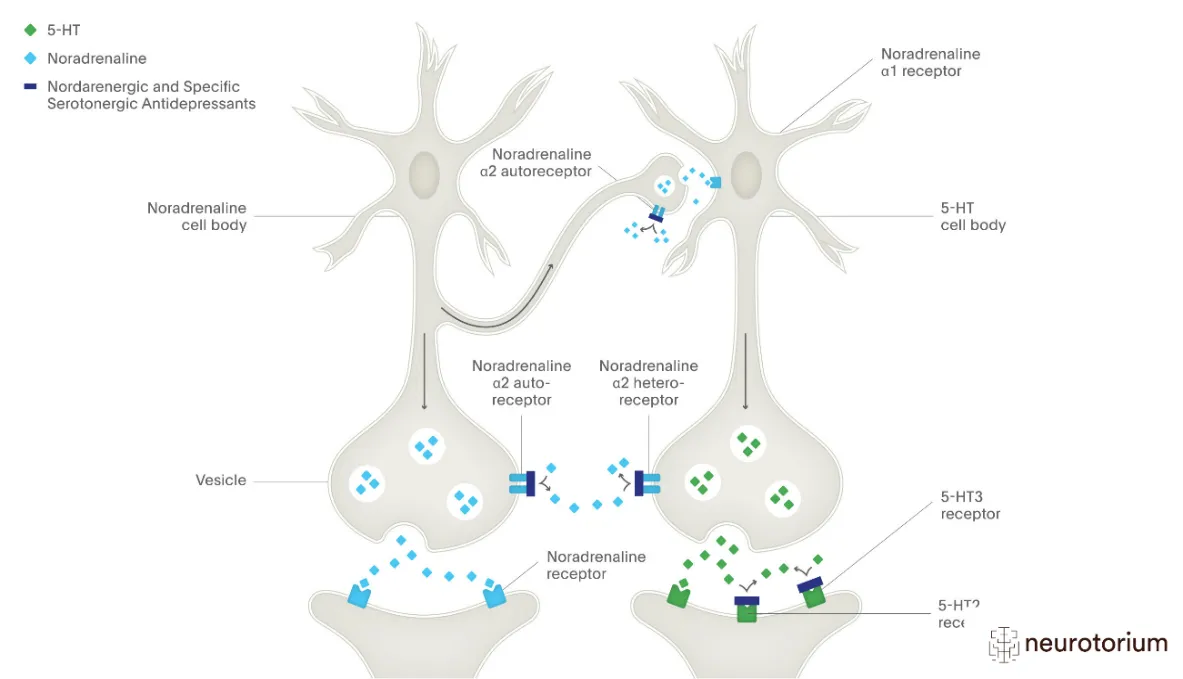
Figure 2: Diagram of serotonergic and noradrenergic pathways.
NEUROBIOLOGY OF DEPRESSION
The serotonergic system is central to mood regulation, cognition, and emotional processing. Dysregulation of serotonin (5‑HT) signaling is strongly implicated in major depressive disorder (MDD). Serotonin receptors, particularly 5‑HT1A and 5‑HT2A, play critical roles in mediating antidepressant responses. Agonism at 5‑HT1A receptors enhances serotonergic neurotransmission, while antagonism at 5‑HT2A receptors reduces hyperactivity associated with anxiety and depression (31,32).
The serotonin transporter (SERT) regulates synaptic serotonin levels. Genetic polymorphisms such as 5‑HTTLPR and epigenetic modifications of the SLC6A4 gene influence susceptibility to depression and treatment outcomes (33). SSRIs act by inhibiting SERT, prolonging serotonin availability in the synaptic cleft, thereby restoring mood balance (34). Recent studies highlight that SERT dysfunction not only alters neurotransmission but also interacts with stress pathways, amplifying vulnerability to depression (35).
Table 3. Key Components of Serotonergic Pathways in Depression
|
Component |
Function |
Dysregulation in Depression |
Reference |
|
5‑HT1A receptor |
Autoreceptor, regulates serotonin release |
Reduced activity, impaired feedback |
(31) |
|
5‑HT2A receptor |
Postsynaptic receptor, modulates mood |
Hyperactivity linked to anxiety/depression |
(32) |
|
SERT |
Serotonin reuptake transporter |
Genetic polymorphisms, epigenetic changes |
(33,34,35) |
Noradrenaline (norepinephrine) contributes to arousal, attention, and stress response. The noradrenaline transporter (NET) terminates noradrenergic signaling by reuptake into presynaptic neurons. Structural studies using cryo‑EM have revealed how NET interacts with antidepressants, providing mechanistic insights into drug binding and inhibition (36,37).
Adrenergic receptors, particularly α2‑adrenoceptors, modulate presynaptic release of norepinephrine. Antagonism of α2‑adrenoceptors enhances noradrenergic tone, producing antidepressant effects (38). SNRIs act by inhibiting both SERT and NET, thereby amplifying serotonergic and noradrenergic signaling simultaneously (39). Dysfunction in noradrenergic circuits contributes to impaired stress resilience and anhedonia, core features of depression (40).
Table 4. Noradrenergic Pathways in Depression
|
Component |
Function |
Dysregulation |
Reference |
|
NET |
Reuptake of norepinephrine |
Altered transporter function, drug binding |
(36,37) |
|
α2‑adrenoceptor |
Presynaptic inhibition |
Overactivity reduces noradrenaline release |
(38) |
|
Adrenergic signaling |
Stress response, arousal |
Impaired resilience, anhedonia |
(39,40) |
Evidence suggests strong synergistic interactions between serotonergic and noradrenergic systems. Electrophysiological studies demonstrate that serotonin modulates noradrenergic firing rates, while norepinephrine influences serotonergic tone (41). This cross‑talk underlies the superior efficacy of dual‑acting antidepressants compared to single‑target agents (42).
Clinical correlates show that patients with combined serotonergic and noradrenergic dysfunction exhibit more severe depressive phenotypes, including cognitive impairment and treatment resistance (43). Thus, integrated modulation of both systems is essential for robust antidepressant responses.
Traditional antidepressants focus on single targets, but multi‑target directed ligands (MTDLs) represent a paradigm shift. These compounds simultaneously modulate multiple neurotransmitter systems, including serotonin, norepinephrine, dopamine, and glutamate (44). Rational drug design strategies now emphasize polypharmacology, aiming to enhance efficacy and reduce side effects (45). Herbal phytochemicals, with their inherent multi‑target actions, align well with this emerging therapeutic approach.
Table 5. Cross‑Talk and Multi‑Target Modulation
|
Mechanism |
Description |
Therapeutic Implication |
Reference |
|
Serotonin → NE |
5‑HT modulates NE firing |
Enhances dual antidepressant efficacy |
(41,42) |
|
NE → Serotonin |
NE influences 5‑HT tone |
Improves mood and cognition |
(43) |
|
Multi‑target ligands |
Polypharmacology across systems |
Higher efficacy, fewer side effects |
(44,45) |
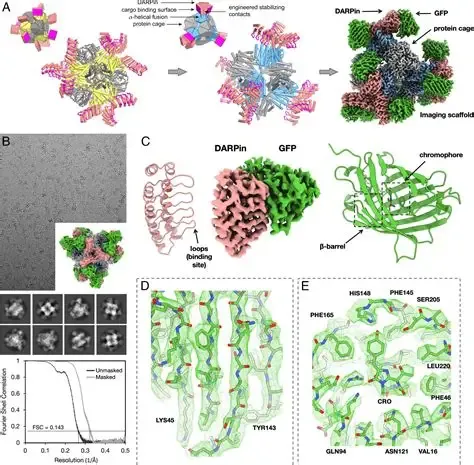
Figure 3: Cryo‑EM structure of NET illustrating antidepressant binding sites.
HERBAL MEDICINES IN NEUROPSYCHIATRY
Table 6. Historical Herbal Remedies for Mood Disorders
|
Tradition |
Herb |
Reported Use |
Reference |
|
Ayurveda |
Ashwagandha |
Stress, anxiety, depression |
(46,47) |
|
TCM |
Bacopa monnieri |
Cognitive and mood enhancement |
(48) |
|
European |
Hypericum perforatum |
Mild‑moderate depression |
(49) |
Table 7. Mechanistic Actions of Phytochemicals
|
Compound |
Class |
Mechanism |
Reference |
|
Quercetin |
Flavonoid |
MAO inhibition, antioxidant |
(50,51) |
|
Harmine |
Alkaloid |
5‑HT receptor modulation |
(52) |
|
Lupeol |
Terpenoid |
Anti‑inflammatory, neuroprotective |
(54) |
|
Ginsenosides |
Terpenoid |
Synaptic plasticity, BDNF upregulation |
(55,57) |
Table 8. Comparative Safety and Efficacy
|
Parameter |
Synthetic Drugs |
Herbal Medicines |
Reference |
|
Onset of action |
2–6 weeks |
Potentially faster |
(58,59) |
|
Side effects |
Sexual dysfunction, insomnia |
Generally fewer |
(60,61) |
|
Cultural acceptance |
Moderate |
High |
(62) |
|
Research status |
Extensive RCTs |
Growing preclinical/clinical evidence |
(63) |
Figure 4: Gut–brain axis illustration showing phytochemical modulation

Figure 5: Botanical illustration of Carissa spinarum shrub.
Table 9. Phytochemical Constituents of Carissa spinarum Root Bark
|
Compound |
Class |
Reported Activity |
Reference |
|
Carissin |
Alkaloid |
CNS stimulation, antidepressant-like |
(72) |
|
Lupeol |
Triterpenoid |
Anti-inflammatory, cardioprotective |
(73) |
|
Quercetin |
Flavonoid |
Antioxidant, MAO inhibition |
(74) |
|
β-sitosterol |
Sterol |
Adaptogenic, neuroprotective |
(75) |
Table 10. Pharmacological Activities of Carissa spinarum
|
Activity |
Mechanism |
Evidence |
Reference |
|
Anti-inflammatory |
Inhibits leukocyte migration |
Animal models |
(77,78) |
|
Cardiotonic |
Enhances myocardial contractility |
Preclinical studies |
(76) |
|
Neuroactive |
Modulates 5‑HT and NA pathways |
Rodent models |
(75) |
|
Antioxidant |
Free radical scavenging |
In vitro assays |
(74,76) |
|
Hepatoprotective |
Protects against toxins |
Animal studies |
(72) |
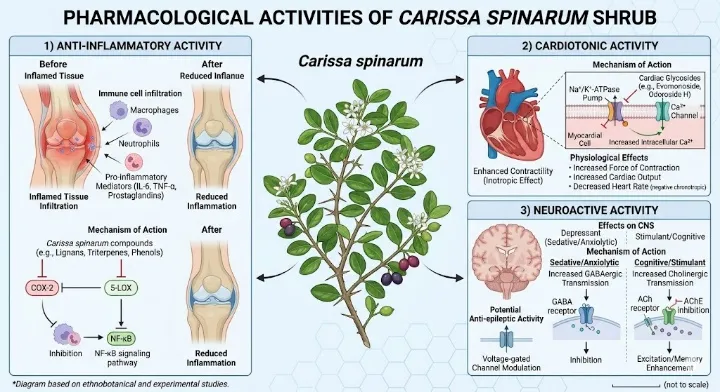
Figure 6: Diagram showing pharmacological activities (anti‑inflammatory, cardiotonic, neuroactive).
EXPERIMENTAL MODELS IN ANTIDEPRESSANT RESEARCH
Rodents, particularly Wistar rats, are widely used in neuropsychiatric research due to their genetic stability, reproducibility, and behavioral responsiveness (79). Their consistent performance in behavioral paradigms makes them ideal for evaluating antidepressant‑like activity. Wistar rats exhibit stress‑induced behavioral changes that closely mimic human depressive phenotypes, including despair, anhedonia, and cognitive impairment (80, 81).
Table 11. Behavioral Paradigms in Wistar Rats
|
Paradigm |
Purpose |
Antidepressant Response |
Reference |
|
TST |
Screens antidepressants |
Reduced immobility |
(82) |
|
OFT |
Differentiates stimulant vs antidepressant |
Normal locomotion |
(83) |
|
SPT |
Measures anhedonia |
Increased sucrose preference |
(84) |
Quantification of serotonin (5‑HT) and norepinephrine (NA) levels in brain tissue, along with receptor binding assays, provides mechanistic insights (85). Techniques include HPLC with electrochemical detection, immunohistochemistry, and receptor autoradiography (86). These assays confirm whether behavioral changes correspond to neurochemical modulation.
Rodent models provide predictive validity for human depression. Many antidepressants currently in use were first validated in FST and TST paradigms (87). While limitations exist—such as differences in stress resilience and cognitive complexity—rodent models remain indispensable for preclinical screening (88).
EVIDENCE OF ANTIDEPRESSANT‑LIKE EFFECTS OF CARISSA SPINARUM
Studies on Carissa spinarum root bark extract demonstrate dose‑dependent antidepressant‑like activity in Wistar rats (89). Solvent fractions significantly reduced immobility in TST, suggesting serotonergic and noradrenergic involvement (90).
Biochemical assays revealed increased serotonin and norepinephrine levels in hippocampal and cortical regions. Extracts modulated 5‑HT1A and α2‑adrenoceptors, supporting dual pathway involvement (93).
When compared to fluoxetine (SSRI) and venlafaxine (SNRI), C. spinarum extract showed similar efficacy in reducing immobility and restoring sucrose preference (90,91). Its multi‑target phytochemical profile suggests potential advantages over single‑target synthetic drugs.
Table 12. Evidence of Carissa spinarum Extract in Rats
|
Dose |
Behavioral Outcome |
Neurochemical Effect |
Reference |
|
Low |
Mild immobility reduction |
Slight 5‑HT increase |
(89) |
|
Moderate |
Significant immobility reduction, ↑ sucrose preference |
↑ 5‑HT, ↑ NA |
(90,91) |
|
High |
Comparable to fluoxetine/venlafaxine |
Strong dual modulation |
(92,93) |
Phytochemicals in Carissa spinarum root bark exert antidepressant‑like effects by modulating serotonergic neurotransmission. Evidence suggests agonism at 5‑HT1A receptors enhances serotonergic tone, while antagonism at 5‑HT2A receptors reduces hyperactivity linked to anxiety (94). Extracts also inhibit the serotonin transporter (SERT), prolonging serotonin availability in the synaptic cleft (95). This dual modulation mirrors the mechanism of SSRIs but with broader receptor activity due to phytochemical diversity (96).
Noradrenergic pathways are influenced by NET inhibition, which increases synaptic norepinephrine levels (97). Phytochemicals such as triterpenoids and alkaloids interact with α2‑adrenoceptors, enhancing presynaptic release of norepinephrine (98). This mechanism resembles SNRIs but may provide additional cardioprotective effects due to the presence of sterols and flavonoids (99).
Unlike synthetic drugs, phytochemicals act in a polypharmacological manner, simultaneously modulating serotonergic and noradrenergic systems. Quercetin, lupeol, and β‑sitosterol collectively enhance neurotransmission, reduce oxidative stress, and improve synaptic plasticity (100). This synergy may explain the comparable efficacy of C. spinarum extract to fluoxetine and venlafaxine in rodent models (101).
Emerging evidence suggests involvement of dopaminergic and GABAergic systems. Flavonoids modulate dopamine turnover, while alkaloids enhance GABAergic inhibition, contributing to anxiolytic and mood‑stabilizing effects (102,103). These secondary pathways highlight the broader neuroactive potential of C. spinarum beyond monoaminergic modulation.
Table 13. Mechanistic Pathways of Carissa spinarum Extract
|
Pathway |
Mechanism |
Phytochemicals |
Reference |
|
Serotonergic |
5‑HT1A agonism, 5‑HT2A antagonism, SERT inhibition |
Quercetin, alkaloids |
(94–96) |
|
Noradrenergic |
NET inhibition, α2‑adrenoceptor modulation |
Lupeol, sterols |
(97–99) |
|
Synergistic |
Dual modulation, antioxidant support |
Quercetin, β‑sitosterol |
(100,101) |
|
Secondary |
Dopaminergic turnover, GABAergic inhibition |
Flavonoids, alkaloids |
(102,103) |
SAFETY AND TOXICOLOGICAL CONSIDERATIONS
Preclinical studies show that C. spinarum root bark extract is generally safe at therapeutic doses. Acute toxicity assays in rodents revealed no mortality up to 2000 mg/kg, while sub‑chronic administration showed mild hepatic enzyme elevation at very high doses (104). Histopathological analysis confirmed absence of major organ damage (105).
Teratogenicity studies in mice indicated potential embryotoxic effects at supra‑therapeutic doses, suggesting caution during pregnancy (106). Reproductive safety data remain limited, but preliminary findings recommend avoiding use in pregnant and lactating women until further validation (107).
Variability in phytochemical composition across regions necessitates standardization of extracts. HPLC fingerprinting and LC‑MS profiling are recommended to ensure consistent alkaloid and flavonoid content (108). Dose optimization is critical to balance efficacy with safety, particularly for long‑term use.
Table 14. Safety and Toxicological Profile of Carissa spinarum
|
Parameter |
Findings |
Reference |
|
Acute toxicity |
Safe up to 2000 mg/kg |
(104) |
|
Sub‑chronic toxicity |
Mild hepatic enzyme elevation |
(105) |
|
Teratogenicity |
Embryotoxic at high doses |
(106) |
|
Reproductive safety |
Limited data, caution advised |
(107) |
|
Standardization |
HPLC/LC‑MS recommended |
(108) |
COMPARATIVE REGULATORY PERSPECTIVES
Table 15. Comparative Regulatory Frameworks for Herbal Antidepressants
|
Agency |
Requirements |
Implications |
Reference |
|
WHO |
Quality assurance, safety, clinical validation |
Global guidance |
(109–111) |
|
EMA |
30 years safe use, standardized extracts |
Traditional use registration |
(112,113) |
|
FDA |
IND approval, clinical trial evidence |
Limited to supplements unless therapeutic claims |
(114,115) |
|
AYUSH |
GMP, pharmacopoeial standards, traditional validation |
Structured Indian framework |
(116,117) |
The World Health Organization (WHO) emphasizes the integration of herbal medicines into national health systems, highlighting their role in primary care and mental health (109). WHO guidelines stress the importance of quality assurance, safety evaluation, and clinical validation before herbal remedies are adopted for psychiatric use (110). Herbal antidepressants must undergo rigorous pharmacovigilance to ensure consistency and minimize risks (111).
The European Medicines Agency (EMA) recognizes herbal medicinal products under the category of “traditional use registration,” requiring at least 30 years of documented safe use, including 15 years within the EU (112). EMA guidelines mandate standardized extracts, validated pharmacological activity, and toxicological safety data (113).
The U.S. Food and Drug Administration (FDA) classifies herbal products as dietary supplements unless they are marketed with therapeutic claims. For psychiatric applications, FDA requires Investigational New Drug (IND) approval and clinical trial evidence (114). This regulatory distinction often limits herbal antidepressants to complementary use rather than mainstream therapy (115).
India’s AYUSH Ministry (Ayurveda, Yoga, Unani, Siddha, Homeopathy) provides a structured framework for herbal psychotherapeutics. AYUSH guidelines emphasize traditional knowledge validation, Good Manufacturing Practices (GMP), and pharmacopoeial standards (116). Herbal antidepressants such as Withania somnifera and Carissa spinarum are evaluated under AYUSH protocols for safety and efficacy (117).
Global harmonization faces challenges due to variability in regulatory definitions, phytochemical composition, and clinical evidence (118). Differences between WHO, EMA, FDA, and AYUSH frameworks create barriers to international acceptance (119). Harmonization requires standardized methodologies, cross‑cultural clinical trials, and unified pharmacopoeial standards (120).
FUTURE DIRECTIONS
Despite promising preclinical evidence, large‑scale randomized controlled trials (RCTs) are essential to confirm efficacy and safety in humans (121). Trials should evaluate dose‑response, long‑term safety, and comparative efficacy with synthetic antidepressants.
Variability in phytochemical content across regions necessitates HPLC fingerprinting, LC‑MS profiling, and DNA barcoding to ensure reproducibility (122). Standardization will enhance regulatory acceptance and clinical reliability.
Herbal antidepressants can be integrated into CAM frameworks, complementing synthetic drugs and psychotherapies (123). This integration aligns with patient preferences for holistic and culturally accepted treatments.
Future research may explore polyherbal formulations targeting multiple neurotransmitter systems (serotonergic, noradrenergic, dopaminergic, GABAergic). Such formulations could provide superior efficacy and reduced side effects compared to single‑compound therapies (120,123).
Table 16. Future Directions in Herbal Antidepressant Research
|
Focus Area |
Description |
Reference |
|
Clinical trials |
Large‑scale RCTs for efficacy and safety |
(121) |
|
Standardization |
HPLC, LC‑MS, DNA barcoding |
(122) |
|
CAM integration |
Complementary use with synthetic drugs |
(123) |
|
Polyherbal formulations |
Multi‑target modulation |
(120,123) |
CONCLUSION
In conclusion, the evaluation of Carissa spinarum root bark extract highlights its promising role as a multi‑target herbal candidate for managing depression. Preclinical evidence demonstrates that its phytochemicals modulate serotonergic and noradrenergic pathways, while also engaging dopaminergic and GABAergic systems, producing robust antidepressant‑like effects in validated rodent models. Beyond efficacy, the plant shows favorable safety in acute and sub‑chronic studies, though teratogenicity data underscore the need for caution in reproductive contexts. Regulatory perspectives from WHO, EMA, FDA, and AYUSH emphasize the importance of standardized extracts, rigorous clinical trials, and harmonized frameworks to ensure global acceptance. Looking forward, the integration of C. spinarum into complementary and alternative medicine, supported by standardized phytochemical profiling and polyherbal formulations, offers a pathway toward safer, culturally accepted, and more effective therapies for depression. This positions C. spinarum not only as a valuable ethnopharmacological resource but also as a potential bridge between traditional knowledge and modern psychopharmacology.
REFERENCES


Swati Jogdand, Rutuja Mule, Pratiksha Jetithor*, Swati Deshmukh, Evaluation Of Antidepressant-Like Effects Of Carissa Spinarum Root Bark Extract In Wistar Rats Via Serotonergic And Noradrenergic Pathways, Int. J. Sci. R. Tech., 2026, 3 (5), 733-749. https://doi.org/10.5281/zenodo.20321154
 10.5281/zenodo.20321154
10.5281/zenodo.20321154