
We use cookies to ensure our website works properly and to personalise your experience. Cookies policy
Environmental Molecular Microbiology Laboratory, Department of Zoology, University of Rajasthan, Jaipur, Rajasthan-302004, India
The accumulation of polyethylene terephthalate (PET) waste and microplastics generated from the waste has increased alarmingly, and mandates the need for sustainable biodegradation strategies. The bacterial PETase enzyme from Ideonella sakaiensis has been studied exclusively for PET biodegradation, along with similar enzymes from other bacterial sources; however, fungal polyester-degrading hydrolases remain underexplored. Therefore, this study investigated the structural and functional similarity between Ideonella sakaiensis PETase and fungal cutinase-like hydrolases employing computational bioinformatics approaches. A fungal hydrolase sequence obtained from UniProtKB was analyzed using BLASTp, multiple sequence alignment, phylogenetic analysis, and structural comparison tools. Conserved catalytic motifs and Ser–His–Asp/Glu catalytic triads were identified among bacterial PETase and fungal enzymes. Functional domain analysis through InterPro Scan confirmed the presence of conserved ?/? hydrolase domains. Furthermore, Structural superimposition using UCSF ChimeraX revealed conservation of the core ?/? hydrolase architecture with a low RMSD value despite divergence in loop regions. The findings suggest that fungal cutinase-like hydrolases possess PETase-associated structural and catalytic characteristics and may play a vital role in future PET biodegradation and microplastic remediation strategies.
Polyethylene terephthalate (PET) is one of the most widely used synthetic plastics around the world, owing to its durability, lightweight nature, and its huge range of usage in packaging, textiles, and beverage containers. However, the continuous buildup of PET waste in both terrestrial and aquatic settings has turned into a major global environmental concern, due to its high resistance to biodegradation (Rajput et al., 2024; Yoshida et al., 2016) Traditional recycling methods via physical or chemical routes are energy-intensive and may produce secondary pollutants; hence, the need of the hour is to develop more sustainable, eco-friendly, and cost-effective biodegradation strategies (Wei and Zimmermann, 2017).
The discovery of polyethylene terephthalate hydrolase (PETase) from Ideonella sakaiensis emerged as a big turning point in plastic biodegradation research. In simple terms, PETase encompasses the capability to break down PET into ecologically sound monomers under mild conditions, indicating its strong potential in plastic recycling through biotechnological approaches (Yoshida et al., 2016). The structural and biochemical analysis of PETase revealed that it belongs to the α/β hydrolase superfamily and possess catalytic Ser–His–Asp triad that resembles cutinase-like esterases (Chen et al., 2018). Further studies revealed that PETase developed specialized groove-like, substrate-binding clefts and rather flexible loop regions that improve PET accessibility as well as catalytic efficiency compared with conventional cutinases (Austin et al., 2018; Joo et al., 2018).
Most currently characterized PET-degrading enzymes come from bacterial sources, while fungal polyester-breaking hydrolases are still less explored, a bit too much in the background. Fungal cutinases and esterases have already been shown to cleave synthetic polyester-type substrates, so it seems fungi could be a meaningful but underused reservoir of PET-degrading biocatalysts (Ronkvist et al., 2009a; Sulaiman et al., 2012). The broad enzymatic diversity found in fungi, together with their potential to cope with varied and extreme environmental conditions, makes these fungal hydrolases good candidates for computational screening and also for structural study.
The recent advances in bioinformatics, protein structure prediction, and computational structural biology have made it possible to spot new enzyme candidates through in silico analysis approaches. Tools for comparative sequence analysis, phylogenetic reconstruction, domain characterization, and three-dimensional structural superimposition are used extensively to reason about functional as well as evolutionary ties among hydrolases responsible for polyester breakdown (Danso et al., 2018). Especially, the availability of highly accurate protein structure prediction tools, like AlphaFold, has really boosted the study and analysis of catalytic layout and structural conservation inside proteins that do not yet have experimentally resolved structures (Jumper et al., 2021).
The present study is focused on evaluating the fungal cutinase-like hydrolases' similarity in both structure and function, with bacterial PETase using one integrated computational approach. The sequences of fungal hydrolases associated with PET degradation-related annotations were retrieved from UniProt and subjected to BLAST-based homology searches, multiple sequence alignment, phylogenetic analysis, InterPro domain characterization, and structural superimposition. The primary goal of the study is to investigate conservation of catalytic residues, α/β hydrolase-associated domains, and active-site architecture between bacterial PETase and fungal enzymes. The findings offer computational proof for the potential of fungal cutinase-like hydrolases in hydrolyzing PET and highlight their possible relevance in future biodegradation and biotechnological applications.
METHODOLOGY
1. Retrieval of PETase and Fungal Hydrolase Sequences
The amino acid sequence for the bacterial polyethylene terephthalate hydrolase (PETase) from Ideonella sakaiensis was retrieved from the UniProt repository (Blum et al., 2021). Keyword-based UniProtKB searches such as “PET degradation enzyme,” “cutinase,” “polyester hydrolase,” and “carboxylesterase” have been used to identify fungal cutinase-like hydrolases associated with PET-degradation related annotations. The fungal sequence retrieved was further used as a query sequence for downstream homology analysis. Protein FASTA sequences were downloaded from the UniProt Knowledgebase (UniProtKB) for subsequent analyses. Aoverview of the computational workflow employed in the present study, including sequence retrieval, homology analysis, multiple sequence alignment, phylogenetic analysis, domain identification, structure retrieval, and structural superimposition, is illustrated in Figure 1.
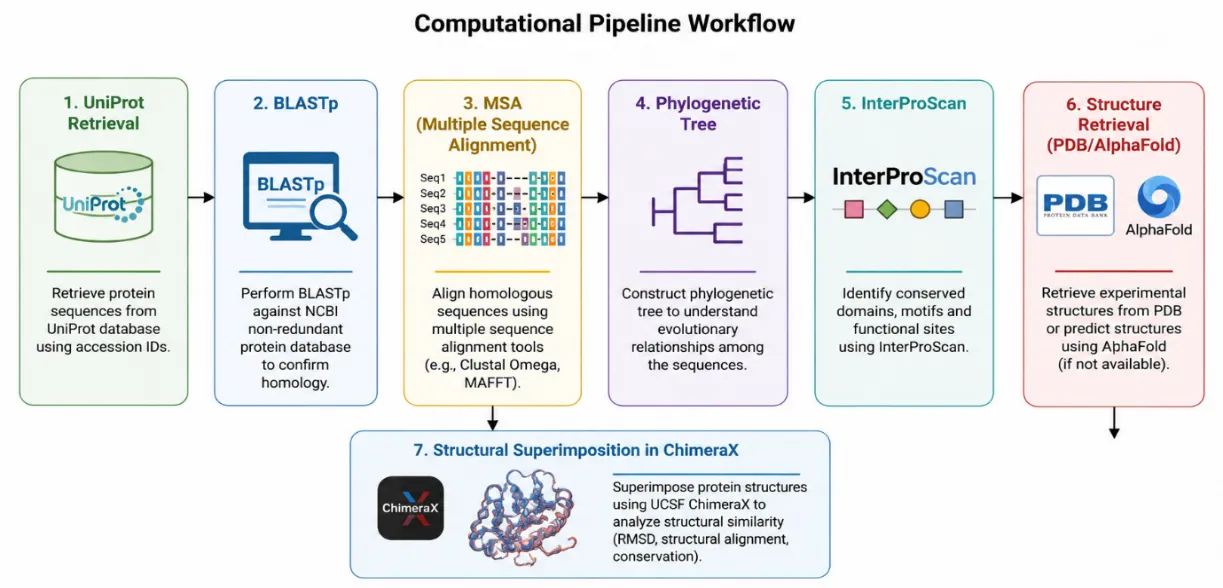
Figure 1. Computational pipeline workflow used for sequence retrieval, homology analysis, phylogenetic analysis, domain identification, structure retrieval, and structural superimposition of fungal hydrolases and bacterial PETase.
2. Sequence Similarity Analysis
The identification of structurally and functionally related hydrolases with the retrieved fungal cutinase-like hydrolase sequence has been done through the similarity searches using the BLASTp algorithm available through the National Center for Biotechnology Information (NCBI) (Altschul et al., 1990). Sequence similarity searches were performed against the non-redundant protein database using default parameters. The protein sequences with size similarity to the retrieved fungal cutinase-like hydrolase were selected for analysis, while fragmented, partial, and uncharacterized proteins were excluded from the study. Further, homologous fungal hydrolases, with conserved catalytic motifs and hydrolase-associated domains, were picked for further investigation.
3. Multiple Sequence Alignment (MSA)
Multiple sequence alignment of PETase and selected fungal hydrolases was performed employing the MUSCLE algorithm implemented in MEGA software(Edgar, 2004; Kumar et al., 2018). The analysis of conserved catalytic residues and motif regions was done manually following alignment generation. The focus was on identifying the catalytic Ser–His–Asp/Glu triad characteristic of α/β hydrolase enzymes. Conserved motifs such as the GxSxG motif associated with catalytic serine residues were analyzed to determine functional conservation among the selected proteins.
4. Phylogenetic Analysis
Phylogenetic relationships among PETase and fungal hydrolase sequences were inferred using the Maximum Likelihood method in MEGA software (Kumar et al., 2018). The evolutionary distances were computed using the Jones–Taylor–Thornton (JTT) substitution model (Jones et al., 1992), and the statistical robustness of the inferred phylogeny was assessed using a bootstrap analysis with 1000 replicates. Further, the resulting phylogenetic tree was subjected to an investigation of evolutionary divergence between bacterial PETase and fungal cutinase-like hydrolases while identifying conserved clustering patterns among related enzymes.
5. Functional Domain and Conserved Fold Analysis
The investigation of functional domain architecture and conserved protein folds was performed using InterProScan (Blum et al., 2021). The presence of α/β hydrolase superfamily domains, cutinase-like domains, esterase-associated motifs, and catalytic residue annotations was analyzed. InterProScan data were used to examine whether fungal hydrolases shared conserved structural properties with bacterial PETase despite low sequence similarity and evolutionary divergence.
6. Three-Dimensional Structural Modeling
Experimentally established PETase structures were procured from the Protein Data Bank (PDB) (Han et al., 2016). Three-dimensional protein models were created using homology modeling or obtained from the AlphaFold Protein Structure Database for fungal hydrolases lacking experimentally determined structures (Jumper et al., 2021). For structural comparison and visualization analysis, predicted structures were downloaded in PDB format.
7. Structural Superimposition and Comparative Structural Analysis
UCSF ChimeraX was used to perform structural superimposition between PETase and fungal hydrolases (Pettersen et al., 2021). MatchMaker tool with default parameters was applied for pairwise structural alignment. The structural similarity between aligned protein regions was estimated using Root Mean Square Deviation (RMSD) values. Structural overlays were used to analyze the catalytic residue location and conserved α/β hydrolase core architecture. Stick representations of the catalytic residues corresponding to the Ser–His–Asp/Glu triad were used to compare the active-site shape of fungal hydrolases and PETase.
8. Visualization and Figure Preparation
Publication-quality structural figures were generated using UCSF ChimeraX. Cartoon representations were used to visualize overall protein fold conservation, while catalytic residues were highlighted using color-coded stick representations. Phylogenetic trees and multiple sequence alignment figures were exported for comparative analysis and graphical representation.
9. Data Interpretation
Comparative analyses were conducted to evaluate the extent of sequence conservation, catalytic residue preservation, α/β hydrolase fold similarity, and structural overlap between bacterial PETase and fungal cutinase-like enzymes. The combined sequence, phylogenetic, domain, and structural analyses were used to infer potential functional similarity and evolutionary conservation of polyester-hydrolyzing mechanisms among fungal hydrolases.
RESULTS AND DISCUSSION
1. Identification of Fungal Cutinase-Like Hydrolases
A fungal cutinase-like hydrolase associated with PET degradation enzyme annotations i.e., Cutinase 1 (EC:3.1.1.74) of 230 Amino acids (AA) from Fusarium vanettenii (Neocosmospora pisi) (UniProt id P00590) was identified through UniProtKB searches. The fungal strain selected for the study owing to its potential to hydrolyze an array of substrates, including soluble esters and insoluble triglycerides, indicating a strong polyester-degradation candidate (Chen et al., 2008; Roussel et al., 2014). Previous studies have also assessed its ability to hydrolyze and alter the surface properties of synthetic polymers and textile fibers such as poly (ethylene terephthalate) (PET), polyamide 6,6, polyester, and acrylic fibers, supporting its significance in PET biodegradation and microplastic remediation research (Araújo et al., 2007). Subsequent BLASTp analysis revealed multiple homologous fungal hydrolases exhibiting significant sequence similarity and conservation of catalytic motifs characteristic of α/β hydrolase enzymes. Furthermore, the selection of partial proteins, hypothetical proteins, and proteins having significant size differences has not been made. Hence, a total of three proteins have been selected, i.e., Cutinase 3 (231 AA) from Dactylonectria estremocensis (KAH7120312.1), Carbohydrate esterase family 5 (233 AA) from Fusarium vanettenii (XP_003042524.1), and cutinase 3 (231 AA) from Fusarium austroafricanum (KAF4452592.1). For consistency throughout the study, Cutinase 3 from Dactylonectria estremocensis was designated as Cutinase 3D, Carbohydrate esterase family 5 from Fusarium vanettenii was designated as CarbEsterase, Cutinase 3 from Fusarium austroafricanum was designated as Cutinase 3F, and Cutinase 1 from Fusarium vanettenii was designated as Cutinase 1. The bacterial polyethylene terephthalate hydrolase (PETase) from Ideonella sakaiensis was referred to as IsPETase throughout the study.
The retrieved fungal proteins were mainly annotated as cutinases, esterases, and carboxylesterases, indicating their potential in hydrolysing the ester bond. Similar hydrolase annotations have previously been associated with microbial polyester-degrading enzymes identified from environmental and metagenomic datasets (Danso et al., 2018; Wei and Zimmermann, 2017). Furthermore, the identification of fungal homologs exhibiting conserved catalytic domains is essential because polyester-degrading enzymes from fungal origin remain comparatively underexplored as compared with bacterial PETases (Ronkvist et al., 2009b)
2. Multiple Sequence Alignment and Catalytic Residue Conservation
Multiple sequence alignment was performed using MUSCLE, demonstrating conservation of catalytic residues among PETase and the selected fungal hydrolases. The GxSxG motif associated with α/β hydrolase catalytic sites has been identified, and hence, the conserved serine residues were also identified within this motif (Figure 2).
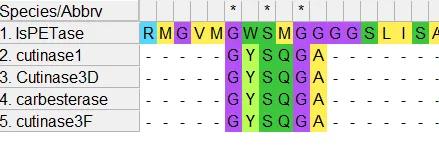
Figure 2: Multiple sequence alignment showing the conserved GxSxG catalytic motif among bacterial PETase and fungal cutinase-like hydrolases. The conserved serine residue within the motif, associated with catalytic activity of α/β hydrolases, is highlighted.
Alongside the catalytic serine, conserved histidine, and aspartate/glutamate residues, were noticed through the fungal sequences, forming the classical Ser–His–Asp/Glu catalytic triad, which is characteristic of cutinase-like hydrolases (Figure 3). The serine residue was positioned at residue 136 in fungal cutinase 1, and in bacterial PETase was located at position 160. Likewise, in fungal cutinase 1, histidine was identified at residue 204, and aspartate was found at 191. In bacterial PETase, histidine and aspartate were identified at 237 and 206, respectively.
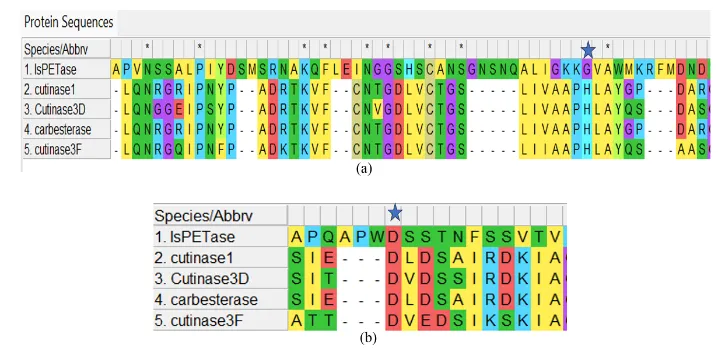
Figure 3: a. Multiple sequence alignment generated using MUSCLE showing conservation of the catalytic histidine residue among fungal cutinase-like hydrolases and bacterial PETase. The catalytic histidine residue is conserved despite differences in sequence position between fungal enzymes (~230 aa) and bacterial PETase (~290 aa), reflecting variation in protein length and sequence architecture. b. Multiple sequence alignment showing conservation of the catalytic aspartate/glutamate residue among fungal cutinase-like hydrolases and bacterial PETase. Although the catalytic residue occurs at different sequence positions due to variation in protein size, its conservation supports preservation of the catalytic triad characteristic of α/β hydrolases.
Despite overall sequence divergence between bacterial PETase and fungal enzymes, the catalytic residues are conserved, indicating the preservation of the hydrolysis mechanism. Previous research work on engineered polyester hydrolases and PETase also paid crucial attention to the conservation of the catalytic triad for efficient ester bond cleavage (Austin et al., 2018; Joo et al., 2018). (Han et al., 2017) reported a similar catalytic architecture in PETase and cutinase-related polyester hydrolases. The presence of this conserved Ser–His–Asp catalytic triad is considered a hallmark feature of cutinase-like polyester hydrolases and plays a vital role in ester bond hydrolysis (Ronkvist et al., 2009).
3. Phylogenetic Analysis
The phylogenetic analysis revealed a distinct monophyletic cluster comprising fungal hydrolases, including cutinase1, carbesterase, Cutinase3D, and cutinase3F. The bootstrap value of 99 indicated the close clustering of cutinase1 and carbesterase, suggesting a strong evolutionary relatedness and high sequence similarity (Figure 4).

Figure 4: Phylogenetic analysis of bacterial PETase and fungal cutinase-like hydrolases performed using the Maximum Likelihood method in MEGA X. The phylogenetic tree demonstrates evolutionary divergence between bacterial PETase and fungal hydrolases while showing conserved clustering among fungal enzymes. Bootstrap values indicate the reliability of branching patterns.
The bacterial PETase was positioned as an outgroup showing branch divergence, while fungal hydrolases formed a separate clade from bacterial PETase. The similar phylogenetic separation between PETase and cutinase-like enzymes has been observed previously in evolutionary analyses of polyester-degrading hydrolases (Danso et al., 2018; Wei and Zimmermann, 2017). This clearly indicates the evolutionary divergence between bacterial PETase and conventional fungal cutinase-like enzymes. However, despite phylogenetic divergence, the presence of the catalytic triad and structural motifs suggests functional conservation of hydrolytic activity.
Previous studies also suggested the evolution of PETase through specialized structural adaptations from ancestral cutinase scaffolds (Han et al., 2016). Also, the comparative evolutionary analysis has also suggested the emergence of PETase-like enzymes through functional optimization of pre-existing cutinase-like hydrolases, comprising the ability to interact with polyester substrates (Sulaiman et al., 2012).
4. InterProScan Domain and Fold Analysis
InterProScan analysis confirmed the presence of α/β hydrolase fold-associated domains in both bacterial PETase and the fungal hydrolases. The persistence of conserved cutinase-like domains and esterase-related signatures has been observed across multiple fungal sequences (Figure 5)



Figure 5: Functional domain analysis using InterProScan showing conserved α/β hydrolase domains in bacterial and fungal polyester-degrading enzymes. (a) Bacterial PETase from Ideonella sakaiensis, (b) Cutinase 1 from Fusarium vanettenii, (c) fungal family carbohydrate esterase, (d) Cutinase 3 from Dactylonectria estremocensis, and (e) Cutinase 3 from Fusarium austroafricanum. All proteins exhibited conserved α/β hydrolase-associated domains, supporting structural and functional similarity among PET-degrading hydrolases.
The identification of α/β hydrolase architecture is highly crucial because PETase belongs to the cutinase-like α/β hydrolase superfamily. The Structural characterization studies of PETase and related polyester hydrolases have demonstrated the presence of conserved α/β hydrolase folds despite taxonomic diversity (Austin et al., 2018; Joo et al., 2018). The presence of similar structural domains in selected fungal enzymes supports the hypothesis that these proteins may possess analogous catalytic mechanisms despite taxonomic divergence. The presence of similar α/β hydrolase-associated domains has also been reported in multiple experimentally characterized polyester-degrading enzymes (Han et al., 2017; Ronkvist et al., 2009).
5. Structural Superimposition Analysis
UCSF ChimeraX was used for the three-dimensional structural superimposition between PETase and representative fungal hydrolases, i.e., Cutinase 1. Structural alignment revealed partial conservation of the α/β hydrolase core architecture despite significant divergence in peripheral loop regions (Figure 6).
The fungal hydrolase Cutinase 1 displayed structural overlap with PETase primarily within the catalytic core region, while external loops and surface regions showed conformational differences. Similar structural variation is consistent with previous reports describing specialized substrate-binding adaptations in PETase relative to ancestral cutinases (Han et al., 2017). Structural studies have shown the presence of widened substrate-binding clefts in PETase and flexible loop regions responsible for enhanced PET accessibility compared to conventional cutinases (Sulaiman et al., 2012).
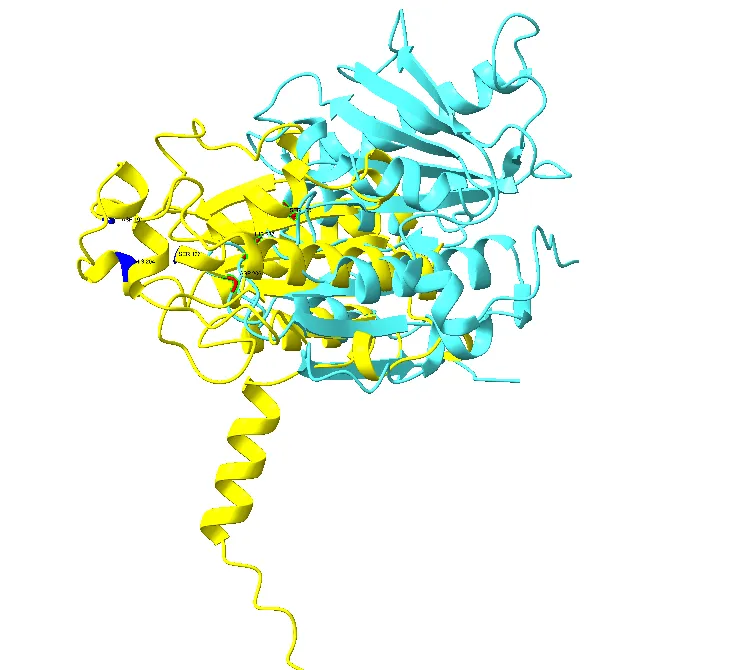
Figure 6. Structural superimposition of bacterial PETase (cyan) and Cutinase 1 from Fusarium vanettenii (yellow) generated using UCSF ChimeraX. The superimposed structures demonstrate conservation of the α/β hydrolase core architecture, with an RMSD value of 1.19 Å indicating significant structural similarity despite evolutionary divergence between bacterial and fungal enzymes.
RMSD analysis demonstrated conservation of localized structural regions despite global divergence. Structural alignment revealed low RMSD values of 1.19 Å within conserved catalytic regions, supporting the presence of a shared hydrolase scaffold.
Despite the variation in the amino acid sequence and protein length, the overall α/β fold architecture remained highly conserved between the bacterial and fungal enzymes, suggesting the preservation of the hydrolase core (Figure 8). Similar α/β hydrolase-associated domains were also identified in other fungal cutinase-like hydrolases through InterPro Scan and were included in the study, suggesting that these enzymes may possess comparable structural organization and catalytic functionality.
The analogous spatial position of catalytic residues Ser–His–Asp/Glu forming the catalytic triad was observed in both PETase and fungal enzymes (Figure 7). This is considered a major determinant of hydrolytic functionality in PETase and cutinase-like enzymes (Austin et al., 2018; Han et al., 2016; Joo et al., 2018). This observation indicates conservation of catalytic geometry and also supports the possibility of mechanistic similarity between the proteins.
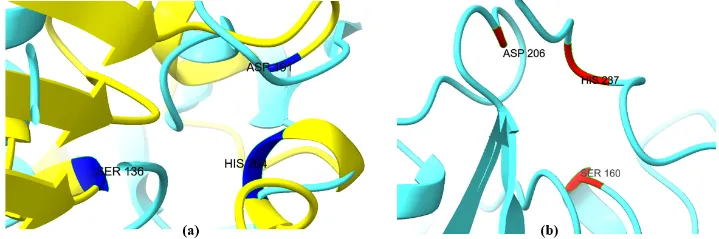
Figure 7. Conserved catalytic triad residues in polyester-degrading hydrolases visualized using UCSF ChimeraX. (a) Catalytic Ser–His–Asp/Glu triad in Cutinase 1 from Fusarium vanettenii and (b) catalytic triad in bacterial PETase. Conserved spatial arrangement of catalytic residues supports mechanistic similarity between fungal cutinase-like hydrolases and bacterial PETase.
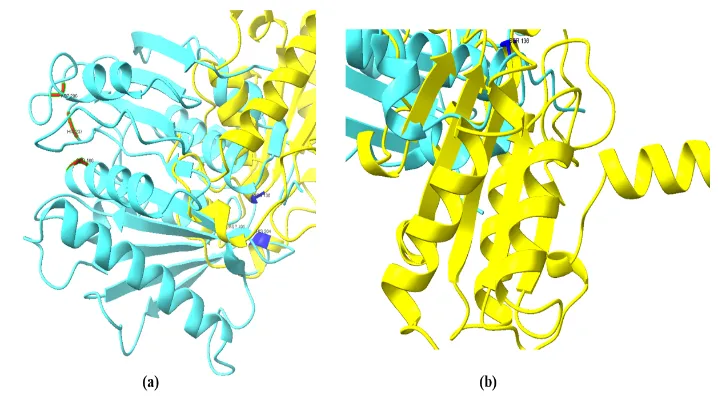
Figure 8. Conserved α/β hydrolase fold architecture observed in (a) bacterial PETase and (b) fungal cutinase visualized using UCSF ChimeraX. Both proteins exhibit the characteristic α/β hydrolase core structure associated with polyester-degrading enzymes, indicating structural conservation despite evolutionary divergence.
CONCLUSION
The present computational study identified fungal cutinase-like hydrolases exhibiting α/β hydrolase-associated domains, conserved catalytic motifs, and structural similarity to bacterial Ideonella sakaiensis PETase. In addition to this, the RMSD analysis clearly showed localized structural region conservation with low RMSD values of 1.19 Å within catalytic regions, indicating the occurrence of a shared hydrolase scaffold. Although these fungal cutinase-like hydrolases analyzed in the present study have not been experimentally validated for PET degradation, the conservation of catalytic residues, α/β hydrolase fold, and active-site geometry clearly suggests the potential polyester-hydrolyzing capability. Furthermore, the results suggest that fungal cutinase-like hydrolases Cutinase 1 from Fusarium vanettenii may represent promising candidates for future polyester degradation studies and experimental validation. The study further highlights the utility of integrated bioinformatics and structural biology approaches for identifying potential PET-degrading enzymes from underexplored fungal sources.
REFERENCES

Meenakshi Rajput*, Deepika, Renu Kumari, Nupur Mathur, Fungal Cutinase-Like Hydrolases As Prospective PET-Degrading Enzymes: An In Silico Analysis, Int. J. Sci. R. Tech., 2026, 3 (5), 1005-1015. https://doi.org/10.5281/zenodo.20415314
 10.5281/zenodo.20415314
10.5281/zenodo.20415314